


The Orthotylinae and Phylinae (Insecta: Heteroptera: Miridae) are world-wide groups currently containing ~4000 highly host-specific phytophagous species (Fig. 1). They comprise about 40 percent of all described species of the plant bug family Miridae , one of the most speciose lineages of non-holometabolous insects. Arguments for the monophyly of the Orthotylinae and Phylinae have been made on the basis of female genitalia (Slater, 1950), male genitalia (Kelton, 1959; Kerzhner and Konstantinov, 1999), and scent-gland morphology (Cassis, 1995), and from a total evidence perspective (Schuh, 1974, 1976). The monophyly of the two subfamilies is corroborated by preliminary analyses of a partial set of the DNA sequence data for 115 species representing 63 genera collected by Wheeler and Schuh under NSF grant DEB-9726587 (see Results of Prior Support). Arguments for sister-group status of the two taxa have also been made (Reuter, 1905; Wagner, 1955; Leston, 1961; Schuh, 1974, 1976).

Comparison of the current classification of the Orthotylinae and Phylinae (Schuh, 1995; http://research.amnh.org/invertzoo/miridae/; Cassis and Gross, 1995) with the baseline work of Carvalho (1952, 1957-1960) indicates significant changes over the last 3 decades (compare Figs. 2 and 3). In the classification of Carvalho (1952) the Phylinae were diagnosed by a plesio-morphic character (setiform parempodia) widely distributed in the Miridae and other Heteroptera, with the consequent inclusion of Dicyphini, a group now placed outside the orthotyline-phyline clade. Also, Carvalho placed the Pilophorini in the subfamily Orthotylinae, solely on the basis of pretarsal structure, an action that Schuh (1974, 1976, 1991), Cassis (1995), and Kerzhner and Konstantinov (1999) have argued is unsupported by other character data. Moreover, because of his very literal interpretation of parempodial structure, Carvalho misplaced a large number of genera, most of them being Phylinae interpreted as Orthotylinae. Lastly, Carvalho diagnosed several tribes on the basis of myrmecomorphic habitus, a "character" that was ill defined, subject to interpretation, and often conflicted with other characters. Many of these "flaws" in Carvalho's classification were addressed by Schuh (1974, 1984, 1991) and Cassis (1995). The higher classification of the subfamilies is now more stable than was that of Carvalho, with most suprageneric taxa being diagnosed on the basis of apomorphic characters.
Significant knowledge of phylogenetic relationships has also been developed below the tribal level. Schuh (1984) prepared a generic-level phylogeny for the tribes Auricillocorini and Leucophoropterini in the Indo-Pacific and for world Pilophorini (Schuh, 1991). Phylogenetic relationships of generic groups in other tribes have been studied by Stonedahl and Schuh (1986), Henry (1991), and Cassis and Asquith (submitted), among others. Species-level phylogenies exist for such complex genera as Pseudopsallus Van Duzee (Schwartz and Stonedahl, 1986; Stonedahl and Schwartz, 1986, 1988) Atractotomus Fieber (Stonedahl, 1990), and Kirkaldyella Poppius (Cassis and Moulds, 2002), among others.
State of Descriptive TaxonomyThe accumulation of described species over time, by classic biogeographic region, can be seen in Fig. 4. Clearly much of the descriptive work in all regions has occurred in the last 3 decades. Possibly the most remarkable feature of these data is that only a tiny part of the described world fauna is from Australia. The figure further suggests that the fauna of all southern continents is depauperate compared to that of the Holarctic, a subject we will address in detail.
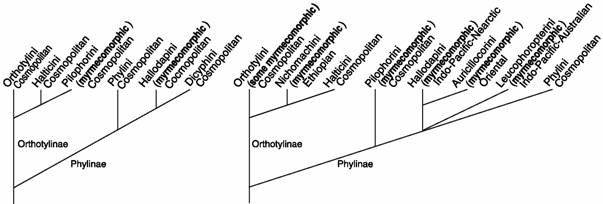
Fig. 2. Carvalho (1952, 1958) Fig. 3. Schuh (1974, 1976, 1984)
Information for Australia indicates that the low species numbers for that region are artificial. For example, as of 1995 CoPI Cassis had borrowed most collections of Australian Miridae to assess the state of taxonomy for the group in the region. These collections contained ~25,000 specimens, of which ~5000 were Orthotylinae and Phylinae, and nearly half of those comprised a few species commonly collected on crop or ruderal plants; ~1000 species of Miridae were represented. Thus, the 180 described species listed by Cassis and Gross (1995) appears to be an abysmal indicator of actual diversity in the Australian Miridae, based on this data source.
PIs Cassis and Schuh began a series of expeditions in 1995 to improve the sample of Australian Miridae. Their efforts have yielded >75,000 specimens from >400 localities (see Fig. 6), most being members of the Orthotylinae and Phylinae. At least 95% of all specimens collected by Cassis and Schuh have documented hosts, 1205 plant vouchers having been identified. Fig. 5 shows the numbers of species of bugs per host, indicating that the sample of Orthotylinae and Phylinae so far collected probably exceeds 1400 species, a very large percentage of which were not present among the ~1000 species found in the pre-existing collections mentioned above. Cassis and Moulds (2002) described in the orthotyline genus Kirkaldyella Poppius 14 species as new from the 15 total that they listed for this Australian group; most specimens came from fieldwork of Cassis and Schuh.
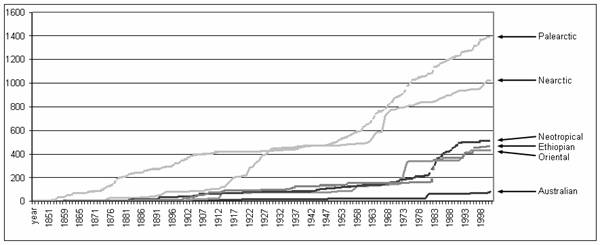
Fig. 4. Accumulation of described species of Orthotylinae and Phylinae over time by continental region.
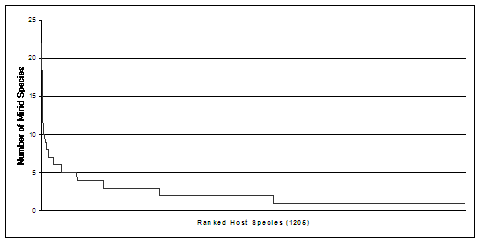
Fig. 5. Count of Australian mirid species by host plant (based on Cassis/Schuh host data).
The sample of Australian Miridae now available for study is immensely greater than what existed 7 years ago. Yet, we believe it still undersamples the fauna because the number of host species recorded underrepresents the total flora, and because the areas sampled by Cassis and Schuh still leave some parts of the continent untouched, as can be seen in Fig. 6. This PBI project offers the opportunity to improve that sample and to describe the fauna comprehensively as part of a larger global study.
Undersampling of the Australian fauna can be further judged by noting that approximately 18,000 species of flowering plants occur in Australia (Costermans, 1994), roughly twice the number found in North America north of Mexico. Within Australia, approximately 3200 plant species occur only in the far southwest (Corrick and Fuhrer, 1996), a region which comprises about 2% of the land area of the continent and is a Mediterranean hotspot. Yet, there are more than 2000 valid species of Miridae known from North America compared to the fewer than 200 recorded for all of Australia (Henry and Wheeler, 1988; Cassis and Gross, 1995; Schuh, 1995). Even a rough correlation between plant bug and plant diversity would predict in excess of 2000 species of Australian Miridae. Data in Fig. 5 suggest that this is almost certainly the case.
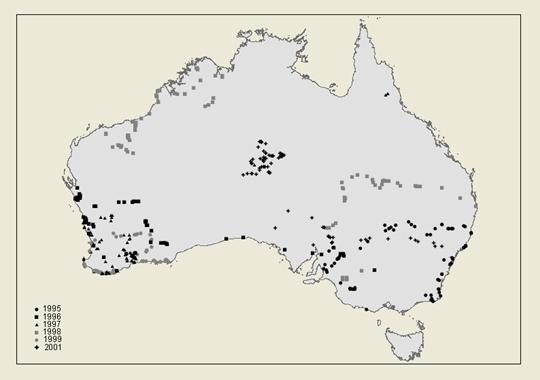
Fig. 6. Australian localities sampled for Miridae by Cassis and Schuh, 1995-2001
A second graphic example of undersampling for the southern fauna is provided by southern Africa, an area with > 23,000 plant species. The Cape Floristic region (fynbos biome) occupies 3.5% of the land area in southern Africa but contains 41% of the plant species, ranking its as one of the botanically most species-rich areas in the world (Cowling and Hilton-Taylor, 1994; Low and Robelo, 1996). Yet, the total numbers of species of Miridae, and more particularly Orthotylinae and Phylinae, currently known from southern Africa is small compared with total plant diversity. Trends of species description suggest that a tremendous amount of diversity remains to be discovered and described. For example, Carvalho et al. (1960) listed 12 species in 12 genera, based solely on the collections of the Lund University Expeditions from the early 1950s. Schuh (1974), who examined a broader range of collections, documented 103 species in 54 genera of Orthotylinae and Phylinae, 81 of the species and 20 of the genera being described as new. A small fraction of the material examined by Schuh was from the far south-west, and virtually none of it was collected during the crucial months of September-November. The remainder of the Ethiopian fauna has been documented in substantial detail, primarily through the works of Linnavuori (e.g., Linnavuori, 1975, 1993, 1994).
Finally, Chile and Argentina are known for their southern biotic connections. PI Schuh conducted field work in the region on 3 separate occasions. Examination of the literature and existing collections indicates that the fauna of the region is not nearly so diverse as those of Australia and southern Africa, but that it is nonetheless crucial from a phylogenetic perspective because of possible biogeographic connections to New Zealand. We have budgeted for additional fieldwork in the region to potentially improve our sample of taxa. The fauna of tropical areas of the Neotropical region has been documented in a very large number of papers by the late J.C.M. Carvalho (see Schuh, 1995). Senior Investigator T. J. Henry has done extensive field work in southern Brazil. These efforts show the Orthotylinae to be much more diverse than the Phylinae, and indicate that significant descriptive work remains to be done (see Table 2).
In summary, although species diversity in the Holarctic appears to be much greater than on the southern continents, the reality appears to be that these fragments of Gondwana are just inadequately studied. Thus, a focal point of this proposal is to collect, document, and describe the southern fauna as part of our effort to monograph the Orthotylinae and Phylinae for the World.
Current Knowledge of Biology, Biogeography, and Biodiversity: Broader Impacts and Non-Systematic Conceptual Issues Biological InformationHost Associations and Specificity: In an effort to assemble information on patterns of host association in the Miridae, Schuh (1995) included ~ 5000 host listings for ~ 2800 species of Miridae. The updated web-based catalog of Miridae, on which Figs. 7 and 8 are based, provides a substantial increase in the number of hosts, with a total of ~ 6200 host records for a total of 3044 species of Miridae, of which ~ 1750 are species of Orthotylinae and Phylinae.
The histograms in Figs. 7 and 8 reveal two strongly supported patterns of host association, based on data for about 30% of described mirid species. First, although most known plant hosts harbor a single mirid species, a significant number of plants serve as hosts for multiple bug species. Second, a preponderance of mirid species appear to be host specific. The 1205 hosts documented by PIs Cassis and Schuh for Australia (see Fig. 5) also portray these patterns.
Host data for the Miridae are collected as a direct result of the bug collecting process. When bug specimens are collected in large numbers from a single plant species, the presumption is that the bug breeds on that plant; if nymphs are present, breeding would seem to be unquestioned. Thus, valuable information on bug behavior/biology can be acquired for the Miridae much more easily than is the case with many groups of phytophagous holometabolous insects, where rearing is required to associate the dissimilar life stages, such as moths and caterpillars.
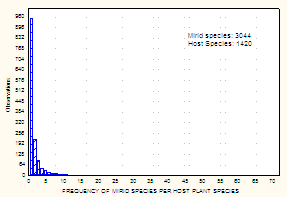
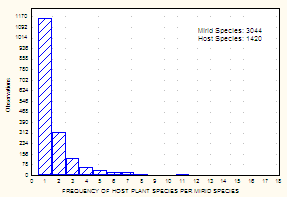
Fig. 7. Data from Schuh (1995 and web update). Fig. 8. Data from Schuh (1995 and web update).
Like many host specific groups, species of Orthotylinae and Phylinae often have more re-stricted distributions than their hosts; this in addition to the fact that a single host species may harbor several species of bugs. These patterns emerge forcefully from data in Figs. 5, 7, and 8. Thus, the amount of information on areas of endemism is greater for bugs than for hosts. For example, Schuh (2000b) showed that in western North America the widely distributed chenopodiaceous hosts Atriplex canescens which harbored 5 species of Megalopsallus Knight and the almost equally widely distributed Sarcobatus vermiculatus which harbored 7 species. Only 2 bug species had distributions anywhere nearly as broad as their host(s).
Myrmecomorphy: Ant-mimicry (myrmecomorphy), occurs widely in the terrestrial Heteroptera, including Miridae (see Fig. 1), and many other groups of insects. Both the nymphs and adults of ant-mimetic Heteroptera are usually of antlike appearance, although the different sexes and life stages may have the appearance of different ant species. At least 4 myrmecomorphic lineages have been documented within the subfamily Phylinae (Schuh, 1986; Fig. 3); the number of independent origins of myrmecomorphy in the Orthotylinae is still an open question, and could range from 2 or 3 to many. Only the type of study described in these pages will resolve this issue. McIver (1987) investigated the dynamics of the relationship between ants and mirids and found that plant bugs provided an excellent experimental system (see also Wheeler, 2001). The phylogenetic knowledge gained from this project will make possible further tests of theories, both historical and ongoing, about the forces that influence the development and maintenance of myrmecomorphy, and whether the primary operators (selective agents) are ants or other organisms. Thus, the results of this study will directly contribute to mimicry theory.
Biogeographic PatternsPlatnick (1991) emphasized the diversity of the faunas in the southern continents, particularly the far south, pointing to the need for further study of these areas to counteract what he called the boreal megafaunal bias in the analysis of animal and plant distributions. We concur with his assessment, and believe that it applies to the Heteroptera (including Miridae) as well as to the spiders documented in detail by Platnick and co-authors (e.g., Forster et al., 1987; Platnick and Forster, 1989; Platnick, 2000, 2002). In support of this contention we refer to the work of Schuh and Slater (1995) which summarized distributions for all families and subfamilies of Heteroptera for the World. Especially in phylogenetically basal clades, we see lineages restricted to the southern continents, suggesting that detailed knowledge of the Gondwana fauna will be essential in the Heteroptera, as in many other groups, if conservation strategies are to preserve lineages as opposed to simply conserving species in terms of raw numbers.
In the classification of Carvalho (1952, 1957-1960) all tribal-level groupings within the Phylinae and Orthotylinae were cosmopolitan (Figs. 2, 3). Schuh (1974, 1976) strengthened arguments for tribal monophyly and produced several restricted distributions (Fig. 3). Schuh (1991) used phylogenetic knowledge of the phyline tribe Pilophorini to show that the most basal lineages in the clade are widespread in the Southern Hemisphere tropics, that lineages arising subsequently are restricted to the Indo-Pacific, and that only the most distal lineages occur in the Holarctic; a similar pattern is seen in the Hallodapini and possibly Leucophoropterini.
Schuh and Stonedahl (1986) and Schuh (1991) used Phylinae and Eccritotarsini to analyze distributions in the Indo-Pacific. Their results corroborate in a rigorous cladistic context general propositions of distribution in the Indo-Pacific as promulgated by Gressitt (1956, 1963), van Balgooy (1971), and others, but which contradict the broad applicability of Wallace's line, a theory with little or no support from cladistic studies in many groups of insects and flowering plants.
In sum, it appears that the Orthotylinae and Phylinae offer substantial potential for in-depth biogeographic analysis and the capability for serving as strong indicators of general patterns of distribution in all continental areas and at a variety of levels in the taxonomic hierarchy, and consequently for different periods of earth history. Only large-scale taxonomic studies of the type described here will lead to persuasive global biogeographic conclusions.
Biodiversity Concepts: Surrogacy and HotspotsPlanetary biodiversity and environmental resources are in substantial decline, particularly as a result of habitat loss and degradation, over-exploitation of species, and the detrimental effects of alien species (Wilson, 1992). Of late, the status of biodiversity is being estimated on a global basis (Myers 1988, Mittermeier et al. 1999, Myers et al. 2000) through the identification of bio-diversity hotspots, which are estimated by an overlay of either centers of high species richness or high endemism and threat information (e.g., human impacts, climate change). Hotspots are being proposed as a credible method for prioritizing conservation efforts. Moreover, their use allows for a broader selection of taxa in identifying areas with high concentrations of biodiversity and makes best use of phylogenetic and collection-based information. The global perspective of the PBI program provides an ideal fit with information requirements of the "hotspot approach".
The "biodiversity crisis" argues for an urgent need to develop biodiversity indicators on a global basis. Because it is operationally impossible to have a single calculus for biodiversity, it is necessary to establish credible "surrogates", whereby one set of organisms is informative of others, particularly in relation to human impacts. The "hotspots approach", as outlined by Myers et al. (2000), predicts that higher plants and vertebrates will serve as surrogates for inverte-brates. However, recent literature (Oliver et al. 1998; Ferrier et al. 1999) indicates that although surrogacy is conceptually attractive, the identification of effective surrogates is elusive. These works suggest that plants and vertebrates are not always effective surrogates for terrestrial invertebrates because their areas of endemism are frequently not coterminous, a phenomenon we have alluded to above under our discussion of Hosts and Biogeographic Patterns.
A more promising approach to identifying biodiversity surrogates is to examine taxa with direct trophic relationships, such as phytophagous insects and their hosts. We argue in this proposal that diversity in the Orthotylinae and Phylinae is strongly correlated with plant diversity, because of the high host specificity in these bugs (Figs. 5, 7, 8). This argument is further sup-ported by work of the Australian Museum Centre for Biodiversity and Conservation Research (Major et al. in press) on Miridae associated with native cypress pine (Callitris glaucophylla).
Conservation International has produced a listing of 25 biodiversity hotspots (Mittermeier et al. 1999). Many of these are found in ecoregions of the southern continents that we argue are grossly under-sampled and inadequately documented for the Orthotylinae and Phylinae. The sampling regime we propose will target already identified hotspots such as the fynbos of South Africa, southwest Australia, and New Caledonia, among others. This study will produce, with PBI funding, documentation to refine the identification of biodiversity hotspots on a global basis.