|
PBI: The Megadiverse, Microdistributed Spider Family Oonopidae
|
|
The Planetary Biodiversity Inventories (PBI) program offers systematists an unparalleled opportunity to "open up" a significant portion of the tree of life to investigation, by a wide array of biologists, on an unprecedented scale. Success in meeting that ambitious goal requires carefully choosing a taxonomic group, assembling an effective team of workers, and demonstrating the broader impacts of the study. We believe that a PBI of the spider family Oonopidae can fulfill those requirements.
|
|
WHY SPIDERS?
|
|
WHY SPIDERS? Spiders are the dominant non-vertebrate predators in almost every terrestrial habitat, making them keystone ecosystem components. They are also extremely diverse, comprising over 2% of all the described species on the planet. Although tackling megadiverse groups like spiders is a daunting task, it is essential that we try, both for science and society, because our current knowledge of biodiversity is highly biased (Platnick, 1999). |
|
Thus, for example, a genus containing three allopatric species within Chile offers much more biogeographic information than does a genus containing only one Chilean species. Not only does the first genus suggest the existence of more areas of endemism, it also offers a potential source of cladistic information: an informative component suggesting that two of those possible areas of endemism share a more recent common history than either does with the third. A single genus containing 20, or even 50, allopatric species within Chile thus has the potential of providing vastly more information on the biotic history of Chile than could even 50 genera of different groups that each contain only a single, widespread Chilean species.
|
|
For policy makers facing the crucial triage situations imposed by today's biodiversity crisis, the same criteria hold. Much has been made of biodiversity "hotspots" -- areas with high species diversity. The now conventional list of such high diversity areas (e.g., Mittermeier et al., 1999) is based almost entirely on vertebrate and plant distributional data. Even if such hotspots turn out to be areas of high diversity for life in general, and even if they turn out to be areas of high endemicity for life in general as well, they offer little potential for making on-the-ground conservation decisions. It is of scant help to a forest manager in Chile to know that the south temperate Nothofagus forests are a biodiversity hotspot. Detailed knowledge of the distribution and interrelationships of a few genera, each with numerous allopatric species inhabiting those forests, on the other hand, could allow narrow areas of endemism to be detected, and used to choose, for preservation attempts, that set of areas that can jointly maximize the phylogenetic diversity we may still be able to save (e.g., Williams et al., 1991).
|
|
There are, of course, many megadiverse groups with microdistributed species, but spiders are unusual among them in several respects. One concerns our ability to deal with existing knowledge. For every megadiverse group, the published systematic literature is astoundingly large, widely scattered, and difficult to access, so that even determining what currently valid species are known in a given taxon, or from a particular region, is a major undertaking. Spiders are one of the few such groups for which our scattered taxonomic knowledge has been completely cataloged in an electronically available form. The World Spider Catalog (Platnick, 2006), which is updated twice a year, provides a complete listing of all available names (and their synonyms), as well as citations to every important published taxonomic treatment of each species, from Linnaeus to date (compilation of the catalog was supported in large part by NSF). The most recent update (Version 7.0) details 39,490 currently valid species, assigned to 3,642 genera and 111 families, and comprises almost 15 megabytes of text. The database conversion of the catalog information by Robert Raven provides Species 2000 and GBIF with their authority files for spiders.
|
|
Spiders are also unusual, among megadiverse groups, in the amount of attention that has been (and is being) focused on their higher-level interrelationships. Arachnologists have long been in the forefront of applying cladistic techniques to the delimitation of higher taxa (e.g., Platnick & Gertsch, 1976; Coddington & Levi, 1991; Hormiga, 1994, 2000; Sierwald, 1997; Griswold et al., 1998, 2005; Arnedo et al., 2004). NSF is currently funding a spider Assembling the Tree of Life (ATOL) award (W. Wheeler, American Museum of Natural History, PI). The numerous participants in that project are assembling a vast set of morphological and molecular data to resolve interfamilial relationships within the order. Four members of our team ( Griswold , Hormiga , Ramírez , and Sierwald ) are active participants in the spider ATOL project), and can thus facilitate a mutually beneficial coordination of effort between the two projects (note, however, that their primary contributions here are their administrative, training, and field expertise, not monographic efforts, so as not to impede their progress on ATOL tasks). |
|
In short, spiders are excellent candidates for a PBI project, because they are megadiverse, often microdistributed, and have a fully cataloged knowledge base, and because their higher-level relationships are currently being studied in an ongoing ATOL project that could interface productively with a PBI project, enabling us to provide a seamless account of both the species-level diversity and the higher-level relationships of a significant chunk of the tree of life.
|
|
WHY OONOPIDS?
|
|
WHY OONOPIDS? Spiders offer a plethora of globally distributed, potential PBI target groups, from the tarantulas (family Theraphosidae), with their macabre public appeal, to the orb-weavers (superfamily Araneoidea) whose intricate webs provide perennially compelling subjects for behaviorists. In choosing a target subgroup, we have followed the same arguments that make spiders compelling overall. Knowledge of currently available collections suggests that hunting spiders in general have smaller ranges than do members of web-building families, and that among hunting spiders, it is the litter-dwelling groups that are likely to have, on average, the smallest ranges of all. These generalizations are based on extensive litter sampling, using mass collecting techniques (Berlese funnels, Winkler extractors, and pitfall traps), conducted by many workers around the globe over the last four decades.
|
|
The spider fauna of litter is dominated by families that were not classically perceived as highly diverse, but that perception is entirely artifactual. Early hand-collecting seldom produced adequate samples of litter-dwelling taxa, which are often quite small in body size and thus easily overlooked by general collectors. Many of the litter-dwelling families belong to the Haplogynae, a relatively basal group of true spiders, and among them, it is unquestionably the Oonopidae (the "dwarf hunting spiders") that dominate most litter samples, both in diversity and abundance. Even the smallest litter sample taken anywhere in the Neotropics, for example, is likely to contain dozens of oonopids, belonging to several different genera and species. In a recent pitfall-trap based survey of spiders in the endangered Atlantic rainforest fragments of Bahia, Brazil, oonopids constituted over 20% of the adult spiders captured, and over 9% of the total species diversity, ranking second only to the jumping spiders (Salticidae) in both abundance and diversity (Dias et al., 2005).
|
|
However, unlike other haplogyne families that are also common in Neotropical litter samples (such as the Tetrablemmidae and Ochyroceratidae), oonopids are well represented in biotas other than tropical rainforest. They occur on all continents (except Antarctica, which has no spiders) and are found in a wide variety of habitats, including foliage, under bark, and even in caves, deserts, bird and termite nests, and the webs of other spiders. In the United States and Canada, where no oonopid genera have yet been the subject of modern revisions and the fauna is therefore significantly understudied, 11 genera and 27 species have been recorded (Ubick, 2005), with the family thus ranking 10th in generic-level diversity, out of the 68 families so far known from north of Mexico (Ubick et al., 2005).
|
|
Perhaps most surprising, though, is that oonopids are also a significant component of the canopy fauna. Canopy habitats are not nearly as well sampled globally as is the litter fauna, but canopy collections from different continents all include oonopids as major components. For example, in the Tanzanian montane forest canopy samples of Sørensen (2004), oonopids were the second most abundant spider family. Although species diversity there was low (only three species reported, from one genus), samples from lowland rainforest canopy in New Guinea (an island from which no oonopids have been described) include at least 11 oonopid species belonging to at least three genera (De Bakker, in litt.). Indeed, oonopids may be unique, among spider families, in being so well represented in both the litter and canopy faunas.
|
|
Given their wide geographic distribution, their surprising microhabitat virtuosity, the fact that adequate litter sampling has been done only in recent decades, and that canopy sampling has been conducted for an even shorter time, it isn't surprising that oonopid diversity has been seriously underestimated. At present, the family contains 459 described species placed in 67 genera (Platnick, 2006). Based on the preliminary data detailed below, we estimate that these described taxa represent only about 20% of the total diversity of the family.
|
|
In short, oonopids are a compelling target for intensive investigation. Among all spiders, they are the group that seems most likely to produce extremely fine-scale information about areas of endemism and their interrelationships, on a global basis, and therefore most likely to be of maximal use to both biogeographers and conservation biologists, once the species are described, their distributions are mapped, and their interrelationships are inferred. Tackling oonopids will fulfill in abundance the basic goals of the Biodiversity Surveys and Inventories program: to discover, describe, and document a hitherto little-explored portion of life on earth. As indicated below, oonopids are also highly unusual spiders, particularly in reproductive morphology, and once their diversity is documented, they are likely to prove a treasure-trove for studies of the evolution of many aspects of comparative biology.
|
|
The Taxa
|
|
As treated in early literature (e.g., Simon, 1893; Dalmas, 1916; Chickering, 1951; Forster, 1956; Hickman, 1979), the group was poorly delimited. Some of the major problems were solved by Forster & Platnick (1985), who were able to corroborate the monophyly of the haplogyne superfamily Dysderoidea, based on two putative morphological synapomorphies: the advancement of the posterior tracheal spiracles to a position just behind the booklung spiracles, and a peculiar specialization of the internal female genitalia, the development of a receptaculum associated with the posterior wall of the bursal cavity.
|
|
Forster and Platnick (1985) showed, primarily by scanning electron microscopy of the tarsal organ (a chemosensory structure found near the tips of the legs and pedipalps), that many of the austral genera previously assigned to the Oonopidae are actually more closely related to Orsolobus (then placed in the Dysderidae) than to true oonopids, and argued that four families of dysderoids should be recognized: the Dysderidae (primarily a Mediterranean group, but with one synanthropic, cosmopolitan species), Segestriidae (a worldwide group of three genera), Orsolobidae (a Gondwanan group, found in Australia, New Zealand, and southern South America, and subsequently discovered in southern Africa by Griswold & Platnick, 1987), and the Oonopidae.
|
|
More recent phylogenetic analyses by Platnick et al. (1991), incorporating new data obtained by scanning electron microscopy of the spinneret spigots through which silk is emitted, confirmed this grouping of families. Resolution within the Dysderoidea varied among their analyses; those authors concluded that "familial relationships within the Dysderoidea (and the monophyly of the Oonopidae) remain uncertain" but favored a sister-group relationship between oonopids and orsolobids.
|
|
Although the monophyly of oonopids, as currently constituted, is not robustly supported, that uncertainty creates no practical problems in delimiting the taxa and specimens to be used in this project (i.e., dysderoids and oonopids, as currently defined, are easily recognizable in collections). Instead, the combination of the work we propose, and that of the spider ATOL project, should allow us to address the issue of oonopid monophyly in detail. We expect that some species currently considered oonopids actually belong elsewhere (e.g., southern African Sulsula may be orsolobids rather than oonopids, and some Neotropical Oonopinus species may be misplaced ochyroceratids), but our project could also conceivably end up demonstrating that oonopids constitute more than a single branch on the dysderoid tree. |
|
Within the family, however, the situation is more stark. The classic, worldwide treatment of Oonopidae by Simon (1893) recognized two subgroups, the "loricati" and "molles." The loricate species are relatively hard-bodied, heavily sclerotized animals in which many of the cephalothoracic sclerites are fused into a single casing, the abdomen typically bears hard dorsal and ventral shields, and the spinnerets are partially surrounded by a chitinous ring (fig. 1). The "molles" forms are weakly sclerotized, soft-bodied animals that lack a dorsal shield on the abdomen (fig. 1). Although often treated as subfamilies (e.g., Roewer, 1942), with the Oonopinae including the soft-bodied taxa and the Gamasomorphinae the loricate ones, it is unlikely that either of those subfamilies is actually monophyletic.
|
|
The same caveats hold for the current generic classification, which can most charitably be described as lamentable. Many species are dumped into relatively large genera that often have few (if any) supporting characters and are surely not monophyletic (this is true even for the type genus, Oonops). On the other hand, more than half of the described genera (35 out of 67) are monotypic and contribute no information on relationships.
|
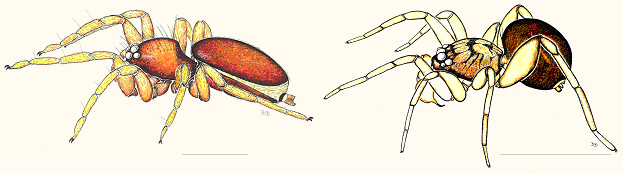
Fig, 1. Left, Gamasomorpha, a loricate oonopid. Right, Orchestina, a soft-bodied oonopid. Scale lines = 1 mm.
|
|
The fact that the current classification of the group is close to meaningless, however, is also not a significant impediment to our project. First, it by no means indicates a scarcity of morphological characters; on the contrary, as indicated by Saaristo (2001: 307): "The family is permeated with peculiar features both in somatic and genitalic characters.
I do not know any other spider family whose members have so many species specific somatic characters as the oonopids." In addition to aiding species recognition, these kinds of morphological specializations make the group ideal for cladistic analysis. Odd somatic features, some of which are sexually dimorphic, include bizarre spines on the head (fig. 2), horns and other extensions on the chelicerae, weird-shaped mouthparts (sometimes producing strange pouches in which the male pedipalps are held in a protected resting position), paired leg spines that can be locked in place via their modified setal bases (fig. 2), hand-like extensions on the posterior coxae, strangely modified pedicels, and many others. Such characters are likely to provide synapomorphies for genera and groups of genera.
|
|
Although haplogynes are typically thought of as having relatively simple genitalia, nothing could be further from the truth in oonopids, which show many features of both the male and female reproductive structures that are apparently without parallels among any other spiders, including the fusion of the cymbium and palpal bulb, enormously swollen palpal patellae; "burned-looking," shriveled and pitch-black palps (fig. 2), etc. Careful studies of the internal female genitalia (e.g., Dumitrescu & Georgescu, 1983, 1987; Burger, Nentwig & Kropf, 2003) have revealed an astonishingly complicated anatomy.
|
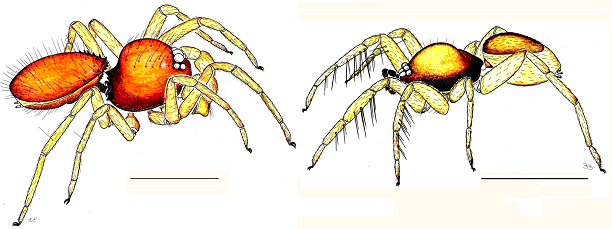
Fig. 2. Left, Xyphinus. Right, Ischnothyreus. Scale lines = 1 mm.
|
|
The severe limitations of the current subfamilial and generic classification pale into insignificance, however, because the proportion of species encompassed is so low. Consider, for example, the Seychelles, where the spider fauna has recently been collected and studied in detail. The classical literature noted only a handful of oonopids from these islands, even if one includes some species originally described just from introduced populations in British greenhouses and only later identified as Seychelle taxa. Studies by Benoit (1979) and Saaristo (1999, 2001, 2002), however, have documented no fewer than 38 species in 20 genera, making oonopids the most diverse of the 40 families known from the Seychelles.
|
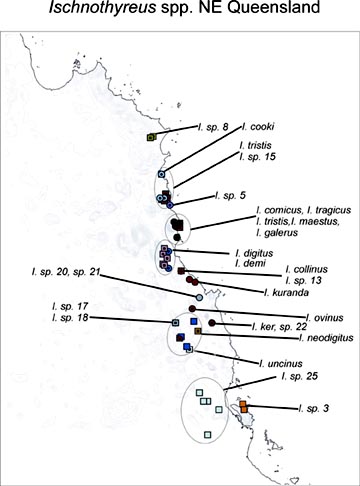 Our extensive preliminary morphological data indicate that those results are not at all atypical. Not a single oonopid species has been described from Madagascar, but the litter fauna there has been collected extensively in recent years, and participant Darrell Ubick , has already separated about 100 species from those collections, in about a dozen (mostly new) genera. Only eight indigenous oonopids have been described from Australia, but participant Mark Harvey's sorting of available Australian collections indicates that the fauna comprises somewhere between 250 and 350 species, belonging to at least 11 genera (of which at least 3 are new). The map on the left shows the distribution of the Ischnothyreus species so far identified from northeastern Queensland; these 25 new species are all microdistributed endemics. In the Ivory Coast, participant Rudy Jocqué's multi-year sampling in the Bossematié forest shows that at least 15 oonopid species co-occur. Although a high proportion of described oonopids are Neotropical, abundant modern litter samples from the New World indicate that the described taxa are again just the icing on the cake, with almost every sample including new species in the Dysderina complex of genera. Even in south temperate Chile, where the dysderoid fauna is dominated by orsolobids and only one true oonopid genus, Unicorn, has been described (Platnick & Brescovit, 1995), Platnick's BS&I-supported fieldwork indicates that at least another six genera are present. Our preliminary surveys of available litter and canopy collections from every continent and most major island groups indicate that the 459 described oonopid species represent only about 20% of the actual diversity of the group. This is a conservative estimate; in Platnick's three recent PEET-supported monographs covering six families of Australasian ground spiders, almost 90% of the species (431 out of 480) had to be newly described.
Our extensive preliminary morphological data indicate that those results are not at all atypical. Not a single oonopid species has been described from Madagascar, but the litter fauna there has been collected extensively in recent years, and participant Darrell Ubick , has already separated about 100 species from those collections, in about a dozen (mostly new) genera. Only eight indigenous oonopids have been described from Australia, but participant Mark Harvey's sorting of available Australian collections indicates that the fauna comprises somewhere between 250 and 350 species, belonging to at least 11 genera (of which at least 3 are new). The map on the left shows the distribution of the Ischnothyreus species so far identified from northeastern Queensland; these 25 new species are all microdistributed endemics. In the Ivory Coast, participant Rudy Jocqué's multi-year sampling in the Bossematié forest shows that at least 15 oonopid species co-occur. Although a high proportion of described oonopids are Neotropical, abundant modern litter samples from the New World indicate that the described taxa are again just the icing on the cake, with almost every sample including new species in the Dysderina complex of genera. Even in south temperate Chile, where the dysderoid fauna is dominated by orsolobids and only one true oonopid genus, Unicorn, has been described (Platnick & Brescovit, 1995), Platnick's BS&I-supported fieldwork indicates that at least another six genera are present. Our preliminary surveys of available litter and canopy collections from every continent and most major island groups indicate that the 459 described oonopid species represent only about 20% of the actual diversity of the group. This is a conservative estimate; in Platnick's three recent PEET-supported monographs covering six families of Australasian ground spiders, almost 90% of the species (431 out of 480) had to be newly described. |
|
Canopy samples are dominated by members of the genus Orchestina (fig. 1), spiders with greatly enlarged femora on the fourth legs that enable them to jump several body lengths. These taxa are of special interest, as their eye pattern suggests that they may be the most basal members of the family. Perhaps unsurprisingly, species of Orchestina also dominate the oonopid fossil record, which consists entirely of amber inclusions in Mesozoic deposits from Lebanon, Myanmar, New Jersey, and Canada as well as the more common Cenozoic ambers (from France, the Baltic area, Mexico, and the Dominican Republic) and subrecent copals (Wunderlich, 2004; Penney, 2006). Although these records include the oldest known amber spider fossils, it is possible that many, most, or even all of the fossils will be assignable to extant species, once our project documents the recent fauna thoroughly.
|
|
|
|
|
|
|
|
|
|
|
| |
|
