|
As treated in early literature (e.g., Simon, 1893; Dalmas, 1916; Chickering, 1951; Forster, 1956; Hickman, 1979), the group was poorly delimited. Some of the major problems were solved by Forster & Platnick (1985), who were able to corroborate the monophyly of the haplogyne superfamily Dysderoidea, based on two putative morphological synapomorphies: the advancement of the posterior tracheal spiracles to a position just behind the booklung spiracles, and a peculiar specialization of the internal female genitalia, the development of a receptaculum associated with the posterior wall of the bursal cavity.
|
|
Forster and Platnick (1985) showed, primarily by scanning electron microscopy of the tarsal organ (a chemosensory structure found near the tips of the legs and pedipalps), that many of the austral genera previously assigned to the Oonopidae are actually more closely related to >Orsolobus (then placed in the Dysderidae) than to true oonopids, and argued that four families of dysderoids should be recognized: the Dysderidae (primarily a Mediterranean group, but with one synanthropic, cosmopolitan species), Segestriidae (a worldwide group of three genera), Orsolobidae (a Gondwanan group, found in Australia, New Zealand, and southern South America, and subsequently discovered in southern Africa by Griswold & Platnick, 1987), and the Oonopidae.
|
|
More recent phylogenetic analyses by Platnick et al. (1991), incorporating new data obtained by scanning electron microscopy of the spinneret spigots through which silk is emitted, confirmed this grouping of families. Resolution within the Dysderoidea varied among their analyses; those authors concluded that "familial relationships within the Dysderoidea (and the monophyly of the Oonopidae) remain uncertain" but favored a sister-group relationship between oonopids and orsolobids.
|
|
Although the monophyly of oonopids, as currently constituted, is not robustly supported, that uncertainty creates no practical problems in delimiting the taxa and specimens to be used in this project (i.e., dysderoids and oonopids, as currently defined, are easily recognizable in collections). Instead, the combination of the work we propose, and that of the spider ATOL project, should allow us to address the issue of oonopid monophyly in detail. We expect that some species currently considered oonopids actually belong elsewhere (e.g., southern African Sulsula may be orsolobids rather than oonopids, and some Neotropical Oonopinus species may be misplaced ochyroceratids), but our project could also conceivably end up demonstrating that oonopids constitute more than a single branch on the dysderoid tree.
|
|
Within the family, however, the situation is more stark. The classic, worldwide treatment of Oonopidae by Simon (1893) recognized two subgroups, the "loricati" and "molles." The loricate species are relatively hard-bodied, heavily sclerotized animals in which many of the cephalothoracic sclerites are fused into a single casing, the abdomen typically bears hard dorsal and ventral shields, and the spinnerets are partially surrounded by a chitinous ring (fig. 1). The "molles" forms are weakly sclerotized, soft-bodied animals that lack a dorsal shield on the abdomen (fig. 1). Although often treated as subfamilies (e.g., Roewer, 1942), with the Oonopinae including the soft-bodied taxa and the Gamasomorphinae the loricate ones, it is unlikely that either of those subfamilies is actually monophyletic.
|
|
The same caveats hold for the current generic classification, which can most charitably be described as lamentable. Many species are dumped into relatively large genera that often have few (if any) supporting characters and are surely not monophyletic (this is true even for the type genus, Oonops). On the other hand, more than half of the described genera (35 out of 67) are monotypic and contribute no information on relationships.
|
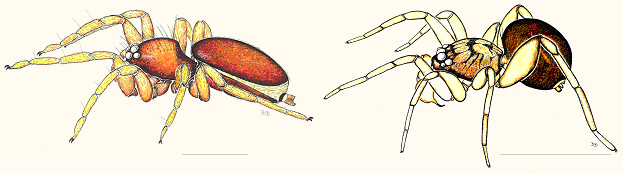
Fig, 1. Left, Gamasomorpha, a loricate oonopid. Right, Orchestina, a soft-bodied oonopid. Scale lines = 1 mm.
|
|
The fact that the current classification of the group is close to meaningless, however, is also not a significant impediment to our project. First, it by no means indicates a scarcity of morphological characters; on the contrary, as indicated by Saaristo (2001: 307): "The family is permeated with peculiar features both in somatic and genitalic characters.
I do not know any other spider family whose members have so many species specific somatic characters as the oonopids." In addition to aiding species recognition, these kinds of morphological specializations make the group ideal for cladistic analysis. Odd somatic features, some of which are sexually dimorphic, include bizarre spines on the head (fig. 2), horns and other extensions on the chelicerae, weird-shaped mouthparts (sometimes producing strange pouches in which the male pedipalps are held in a protected resting position), paired leg spines that can be locked in place via their modified setal bases (fig. 2), hand-like extensions on the posterior coxae, strangely modified pedicels, and many others. Such characters are likely to provide synapomorphies for genera and groups of genera.
|
|
Although haplogynes are typically thought of as having relatively simple genitalia, nothing could be further from the truth in oonopids, which show many features of both the male and female reproductive structures that are apparently without parallels among any other spiders, including the fusion of the cymbium and palpal bulb, enormously swollen palpal patellae; "burned-looking," shriveled and pitch-black palps (fig. 2), etc. Careful studies of the internal female genitalia (e.g., Dumitrescu & Georgescu, 1983, 1987; Burger, Nentwig & Kropf, 2003) have revealed an astonishingly complicated anatomy.
|
|
The severe limitations of the current subfamilial and generic classification pale into insignificance, however, because the proportion of species encompassed is so low. Consider, for example, the Seychelles, where the spider fauna has recently been collected and studied in detail. The classical literature noted only a handful of oonopids from these islands, even if one includes some species originally described just from introduced populations in British greenhouses and only later identified as Seychelle taxa. Studies by Benoit (1979) and Saaristo (1999, 2001, 2002), however, have documented no fewer than 38 species in 20 genera, making oonopids the most diverse of the 40 families known from the Seychelles.
|
Our extensive preliminary morphological data indicate that those results are not at all atypical. Not a single oonopid species has been described from Madagascar, but the litter fauna there has been collected extensively in recent years, and participant Darrell Ubick has already separated about 100 species from those collections, in about a dozen (mostly new) genera. Only eight indigenous oonopids have been described from Australia, but participant Mark Harvey's sorting of available Australian collections indicates that the fauna comprises
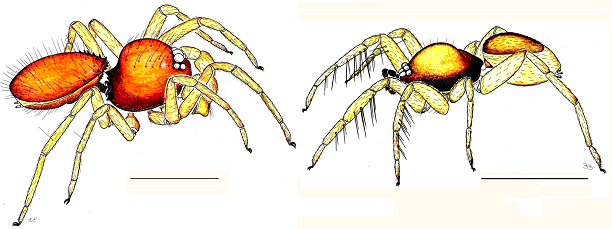
Fig. 2. Left, Xyphinus. Right, Ischnothyreus. Scale lines = 1 mm.
somewhere between 250 and 350 species, belonging to at least 11 genera (of which at least 3 are new).
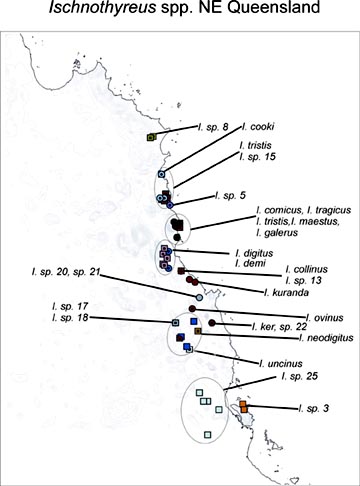 The map on the left shows the distribution of the Ischnothyreus species so far identified from northeastern Queensland; these 25 new species are all microdistributed endemics. In the Ivory Coast, participant Rudy Jocqué's multi-year sampling in the Bossematié forest shows that at least 15 oonopid species co-occur. Although a high proportion of described oonopids are Neotropical, abundant modern litter samples from the New World indicate that the described taxa are again just the icing on the cake, with almost every sample including new species in the Dysderina complex of genera. Even in south temperate Chile, where the dysderoid fauna is dominated by orsolobids and only one true oonopid genus, Unicorn, has been described (Platnick & Brescovit, 1995), Platnick's BS&I-supported fieldwork indicates that at least another six genera are present. Our preliminary surveys of available litter and canopy collections from every continent and most major island groups indicate that the 459 described oonopid species represent only about 20% of the actual diversity of the group. This is a conservative estimate; in Platnick's three recent PEET-supported monographs covering six families of Australasian ground spiders, almost 90% of the species (431 out of 480) had to be newly described.
The map on the left shows the distribution of the Ischnothyreus species so far identified from northeastern Queensland; these 25 new species are all microdistributed endemics. In the Ivory Coast, participant Rudy Jocqué's multi-year sampling in the Bossematié forest shows that at least 15 oonopid species co-occur. Although a high proportion of described oonopids are Neotropical, abundant modern litter samples from the New World indicate that the described taxa are again just the icing on the cake, with almost every sample including new species in the Dysderina complex of genera. Even in south temperate Chile, where the dysderoid fauna is dominated by orsolobids and only one true oonopid genus, Unicorn, has been described (Platnick & Brescovit, 1995), Platnick's BS&I-supported fieldwork indicates that at least another six genera are present. Our preliminary surveys of available litter and canopy collections from every continent and most major island groups indicate that the 459 described oonopid species represent only about 20% of the actual diversity of the group. This is a conservative estimate; in Platnick's three recent PEET-supported monographs covering six families of Australasian ground spiders, almost 90% of the species (431 out of 480) had to be newly described.
|
|
Canopy samples are dominated by members of the genus Orchestina (fig. 1), spiders with greatly enlarged femora on the fourth legs that enable them to jump several body lengths. These taxa are of special interest, as their eye pattern suggests that they may be the most basal members of the family. Perhaps unsurprisingly, species of Orchestina also dominate the oonopid fossil record, which consists entirely of amber inclusions in Mesozoic deposits from Lebanon, Myanmar, New Jersey, and Canada as well as the more common Cenozoic ambers (from France, the Baltic area, Mexico, and the Dominican Republic) and subrecent copals (Wunderlich, 2004; Penney, 2006). Although these records include the oldest known amber spider fossils, it is possible that many, most, or even all of the fossils will be assignable to extant species, once our project documents the recent fauna thoroughly.
|
| |
|
