Interval 3
Interval 3 – Birth of the Modern World

This scene depicts late spring in the Northern Great Plains about 10 million years ago, in what is now southern South Dakota. The Black Hills appear in the background, and a shallow stream with braided sandbars winds through the foreground. Two species of Miocene perissodactyl are present in this picture; the rhino Teleoceras (right) and an extinct horse, Neohipparion (left). Rob Barber\AMNH.
Geologists divide the Cenozoic into two major time periods, the Paleogene and the Neogene. The Paleogene, which includes the Paleocene, Eocene, and Oligocene epochs, is the period in which mammals diversified from relatively small, simple forms into a plethora of diverse animals in the wake of the mass extinction that ended the preceding Cretaceous Period. The continents drifted into more-or-less the configuration that the occupy today and the world’s climate shifted from the relatively warm “greenhouse” state that had prevailed for much of the previous 100 million years to today’s relatively cold “icehouse” conditions. By the end of the Paleogene, the Earth was cooler and drier; rainforests had retreated to the tropics, open woodland and grassland was spreading, and most of the groups of archaic mammals had gone extinct. While individual mammal species from this time might look very strange to our eyes, the major groups that are present today were all present. The beginning of the Miocene epoch, 23 million years ago, is not just the beginning of the Neogene. In a very real sense, it is the beginning of the modern world.
The global trend during the Miocene was towards increasing aridity, caused primarily by global cooling reducing the ability of the atmosphere to absorb moisture. Grasslands underwent a major expansion; forests fragmented and retreated in response to the generally cooler and drier climate overall. Grasses also diversified greatly, co-evolving with large herbivores and grazers. Between 7 and 6 million years ago, there was a sudden expansion of grasses which were able withstand water loss and assimilate carbon dioxide more efficiently. But these grasses were also richer in silica, making them harder to digest. The result was a worldwide extinction of large herbivores.
Too Much of a Mouthful
The success of the grasses lies partly in the their morphology and growth processes, and partly in the diversity of ways that they generate nutrients through photosynthesis. Grasses can be divided into two physiological groups, using two different photosynthetic pathways for carbon fixation, known as C3 and C4. The C4 grasses have a specialized leaf anatomy that particularly adapts them to hot climates and an atmosphere low in carbon dioxide. This is because under conditions of drought, high temperatures and nitrogen or carbon dioxide limitation, 97% of the water taken up by C3 plants is lost through the leaves. In C4 plants this proportion is much lower, giving them an evolutionary edge in a dry environment. The first C4 plants evolved around 25 to 32 million years ago during the Oligocene, probably in response to the development of more open habitat with higher levels of sunlight. By 6 to 7 million years ago, the increasing aridity of the Miocene meant that C4 grasses, with their increased resistance to water stress, were at an advantage compared with C3 grasses. But C4 grasses were also much richer in Silica, making them tougher to eat and digest.
The mammal fauna of Miocene North America was strikingly similar to that of a modern African savannah, but with certain crucial difference. Instead of hippos, there were hippo-like rhinos called Teleoceras, and in place of giraffes there were giant, long-necked camels of the genus Aepycamelus. Tapirs took the place of the African bush pigs and forest hogs. And the role of small to medium-sized browsers and grazers, which in Africa is filled by the antelopes, was taken by a mix of camels, pronghorns, and horses. The Miocene grasslands of North America were dominated by horses; in some fossil localities, as many as 13 different species can be found co-existing in the same place and time.
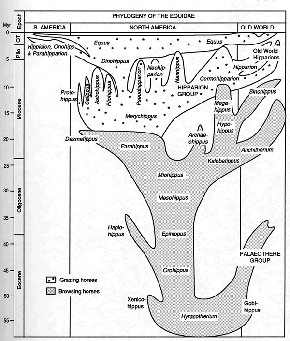
A Different View of Horse Evolution
Traditional views of horse evolution have tended to concentrate on a linear process, from the small, four-toed Hyracotherium to the large, one-toed Equus. However, there is an equally compelling argument that the process involved a series of adaptive radiations of species, with the crucial stage being driven by climate and vegetation during the Miocene. At the end of the Oligocene, horses were still primarily browsers. They had low crowned teeth, suitable for eating leaves and fruit, but there is evidence from microwear studies that they may have started to include some grass in their diet as the open woodland of the Oligocene was gradually replaced by true grassland. Fast-forward to 18 million years ago, and two very different radiations of horses are present. There are still browsing horses, represented in North America by Hypohippus and Archaeohippus. But now there are also true grazing horses, such as Merychippus, exploiting the expanding grasslands. Merychippus has wide molars with flat grinding surfaces and a tough layer of cementum to help cope with a diet of abrasive grasses. By 10 million years ago the browsing horses are all extinct in North America. Now there is a radiation of 8 genera of specialist grazers. But another wave of extinctions towards the end of the Miocene, 6 million years ago, perhaps associated with the spread of C4 left only a single surviving lineage, the ancestor of our modern horses and zebras.
Although the overall trend of the Miocene was a cooling, drying one, there were periods of increased warmth when ungulates diversified. But around 6-7 million years ago the temperature dipped sharply again. This was also the period when the C4 grasses expanded and diversified enormously. Rhinos went extinct in North America, along with the chalicotheres, and the diversity of horses was greatly reduced. By the end of the Miocene, the average global temperature was only 14 degrees Centigrade, compared with over 28 degrees Centigrade at the height of the Eocene. The Earth was about to enter the ice ages.
Coping with Grass
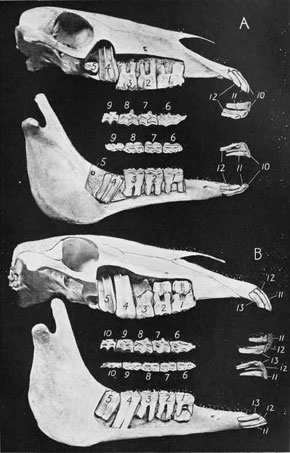
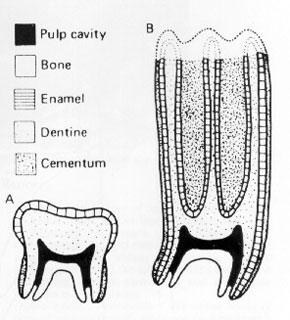
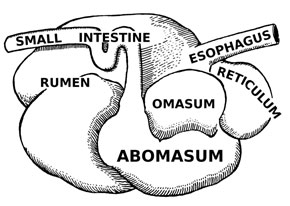
Grass is so abundant today, and supports such large communities of herbivores, that it’s easy to forget what a challenging diet it represents. It’s tough and abrasive – difficult to chew and difficult to digest effectively. Herbivorous mammals cope with grass in a variety of ways. The first is to develop molars and premolars that are capable of withstanding the continuous wear caused by chewing grass. These molars tend to be tall, with enamel that extends well past the gum line, and with hard ridges of enamel exposed on their flat grinding surfaces. These are known as hypsodont teeth and they can be found in almost all mammals that feed on grass. The second is to modify the digestive system. All mammals that eat plants have a flora of microbes that live in their guts – these bacteria release enzymes called cellulases, which break down cellulose and release energy. Plants are rich in cellulose and without the presence of these microbes animals would be unable to digest them effectively. Digesting large volumes of plant material requires large quantities of microbes; herbivorous mammals have evolved to house these microbes by turning their guts into giant fermentation vats. Grass is particularly challenging. One group of mammals, the artiodactyls, have evolved to successfully exploit this resource by developing a multi-chambered stomach, through which food can be passed more than once. This allows for more efficient digestion and a greater range of body sizes. Artiodactyls that do this are called ruminants. By contrast, perissodactyls never evolved this ability and are still dependent on relatively inefficient fermentation in the gut. This may be one reason why today artiodactyls are far more diverse and abundant than perissodactyls (224 species vs. 16 species).
Teleoceras and Aphelops: Two Very Different Rhinos
Rhinos in the modern African savannah exist as a browser-grazer species pair; the white rhinos of the genus Ceratotherium are wide-mouthed grazers, whereas the black rhino Diceros is a browser that uses its beak-shaped lip to pluck leaves and twigs. In Miocene North America there was a similar species-pairing, although the two rhinos concerned were only distant relatives of the modern African species. The aceratherine rhino Aphelops was the browser. The greatly-retracted nasal incision in its skull is evidence for the large muscles needed to operate a prehensile upper lip, and instead of upper incisors it has a horny nipping pad, similar to that of modern rhinos, to help pluck leaves and shoots. By contrast, Teleoceras, a short-legged, hippo-like animal, has high-crowned teeth suitable for grazing and a huge, barrel-shaped body, presumably to support a super-sized gut for digesting tough grasses. Both types of rhino were abundant in the Miocene, but by the early Pliocene both were extinct and rhinos had vanished from North America.
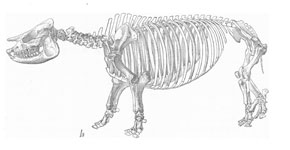
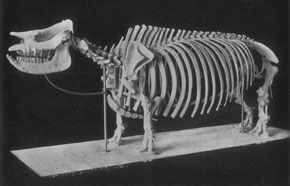
Left: Skeleton of the Miocene browsing rhino Aphelops. AMNH. Right: Mounted skeleton of the Miocene rhino Teleoceras. AMNH.




{kind=link}
{kind=link}
.png){kind=link}